
|
|
|
|
|
|
Shark Info (29.06.1996) |
Author |
|
Intro: |
Shark Info |
|
Hauptartikel: |
Shark Info |
|
Artikel 1: |
Shark Info |
|
Fact Sheet: |
Shark Info |
|
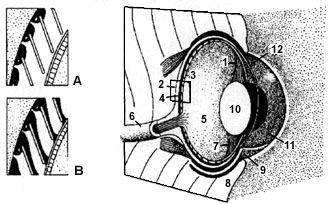
Fact Sheet: Die Augen des HaieHohe Anforderungen an das visuelle System Haie leben sowohl in küstennahen Flachwasserregionen wie auch in der Hochsee. Gewisse Arten wechseln im Laufe ihrer Entwicklung von einem Habitat in das andere, während andere sich an das Leben in der Tiefsee angepasst haben. Die meisten Haie sind sowohl tag-, dämmerungs- als auch nachtaktiv. Die damit verbundenen wechselnden Licht- und Sichtverhältnisse sowie weitere optische Randbedingungen führten zu einem gut entwickelten visuellen System, das sich an die unterschiedlichen Anforderungen der Umwelt angepasst hat. Aufbau des AugesHaiaugen haben mit jenen des Menschen viele Ähnlichkeiten. So kann sich die Pupille durch Öffnen und Schliessen der Intensität des einfallenden Lichtes anpassen eine Fähigkeit, die den Knochenfischen fehlt. Allgemein wird die Brechkraft des Auges durch das Medium, die vordere und hintere Hornhaut und die vordere und hintere Linsenfläche beeinflusst. Da die Hai-Hornhaut (Cornea) unter Wasser auf die Lichtbrechung keinen Einfluss hat, wird die Linse zur optischen Hauptstruktur. Diese ellipsenähnliche, kristalline Linse besitzt einen sehr hohen Lichtbrechungswert (refraktiven Index). Bei Zitronenhaien beträgt er 1.66, was einem Dioptriewert von rund 140 entspricht. Im Vergleich dazu besitzt die menschliche Linse lediglich einen Wert von ca. 20 Dioptrien. Die Augenbewegungen werden von drei Paaren extraocularer Muskeln kontrolliert, welche einerseits schnelle Bewegungen des Auges etwa bei der Verfolgung von Beutetieren andererseits langsamere zur Fokussierung des Bildes auf der Netzhaut (Retina) ermöglichen.
Einige Arten, zum Beispiel der Ammenhai (Ginglymostoma) haben gut entwickelte und mobile Augenlider. Bei anderen wie etwa bei Grauhaien (Carcharinidae) hat sich das untere Lid zu einer Art Nickhaut entwickelt, wie sie bei höheren Wirbeltieren (z.B. bei Adlern) vorhanden ist. Bei Reizungen der augennahen Haut und beim Öffnen des Kiefers etwa beim Angriff auf ein Beutetier schiebt sich die Nickhaut schützend vor das Auge. Beim Weissen Hai fehlt sie; er schützt sich, indem er den Augapfel beim Öffnen des Kiefers gegen hinten dreht. OptikAlle bis anhin untersuchten Netzhäute besitzen beide bekannten Typen lichtsensibler Zellen, nämlich Zäpfchen und Stäbchen. Die Anzahl der Stäbchenzellen überwiegt, was die Sehfähigkeit bei schlechten Lichtverhältnissen erhöht. Herausstechend ist auch der Umstand, dass die Zusammensetzung der lichtempfindlichen Chemikalien in den Stäbchenzellen auf die Wellenlängen des zur Verfügung stehenden Lichtes genau abgestimmt ist. Bei gewissen Haien ändert sich die Zusammensetzung dieser Sehpigmente beim Wechseln des Habitats. Für eine weitere Sensivitätssteigerung sorgt eine reflektierende Schicht hinter der Retina: das sogenannte Tapetum luciduum. Licht, das durch die Sehzellen dringt, wird durch dieses Tapetum im Augenhintergrund reflektiert, was dazu führt, dass die Sehschicht ein zweites Mal gereizt wird. Wird ein Haiauge angestrahlt, leuchtet es wie ein Katzenauge. Das Tapetum luciduum ist eine Struktur, die mit reflektierenden Guaninplättchen bedeckt ist. Diese sind in einem Winkel angeordnet, der das Licht entlang der Stäbchen-Längsachse auf die Retina zurückwirft. Derartige Reflektoren können darauf abgestimmt werden, Licht auf ganz bestimmte Wellenlängenbereiche zu reflektieren. Die Reflektoren von Tiefsee- haien beispielsweise spiegeln das Restlicht, das in tiefe Wasserschichten vordringt, optimal. Haie können Farben erkennen. Davon gehen neuere Forschungen aus, insbesondere beim Zitronenhai. Diese Art verfügt über eine weitere Spezialisierung: In einem bestimmten Abschnitt der Retina hat der Zitronenhai eine erhöhte Anzahl lichtempfindlicher Zellen, den sogenannten visuellen Streifen. Dieser ermöglicht ihm, während dem Patrouillieren knapp über dem Meeresboden sein Aktionsfeld visuell optimal zu erfassen. Auffallend ist, dass Grösse und Lage der Augen bei verschiedenen Arten stark variieren. Wenig aktive, die flachen Gewässer bewohnenden Haie wie zum Beispiel der Stierkopf (Heterodontus) und Ammenhai (Ginglymostoma sp.) haben kleine Augen, deren Durchmesser unter einem Prozent ihrer Körperlänge liegt. Dies lässt den Schluss zu, dass sich solche Arten bei der Nahrungssuche weniger auf den Gesichtssinn als auf andere sensorische Systeme stützen. Dagegen weisen aktive Jäger wie etwa der Drescherhai (Alopias) die in tieferem Wasser operieren, wesentlich grössere Augen auf. Entgegen einer weit verbreiteten Meinung, Haie sähen schlecht, muss heute angenommen werden, dass sie ihren Bedürfnissen entsprechend optimal angepasste Augen haben.
Verwendete Literatur: Bone, Q. & N.B. Marshall (1985). Biologie der Fische. Gustav Fischer Verlag. 154-186 Englische Originalausgabe 1982 bei Blackie & Son Limited, Glasgow. Catt, M., Ernst, W. & C.M. Kemp (1983). The products of photoreversing rhodopsin bleaching by microsecond flashes in the isolated vertebrate retina. Vision Res. 23: 971-982. Graeber, R.C. (1978). Behavioral studies correlated with central nervous systemintegration of vision in sharks. In: Hodgson, E.S., Methewson, R.F. (eds). Sensory biology of sharks, skates, and rays. 195-225. Graf, W.G. & W.J. Brunken (1984). Elasmobranch oculomotor organization: anatomical and theoretical aspects of the phylogenetic development of vestibulo-oculomotor connectivity. J. Comp. Neurol. 222: 569-581. Housley, G.H. & J.C. Montgomery (1984). The structure of the external rectur muscles of the carpet shark Cephaloscyllium isabella. J. Ant. 138: 643-655. Montgomery, J.C. (1990). Sensory physiology. In: Shuttleworth, T.J. (ed). Physiology of Elasmobranch Fishes. 83-88. Veröffentlichung nur mit Quellenangabe: Shark Info |
|||||
|
|
|
|
|
|||||